Dr Jonathan Bujak has been researching the biostratigraphic zonal schemes of the Paleogene for a number of years. His non-exclusive well database documents the Paleogene stratigraphy and absolute ages in 1155 offshore UK and Norwegian wells, including the oceanographic and climatic changes described in these articles.
This, the first of two blogs, documents the interpreted oceanographic and climatic changes that occurred in the North Atlantic region during the Paleogene. Our next blog on this topic will discuss their relevance for energy exploration and carbon capture & storage.
Paleogene Climate and Oceanography
The North Atlantic region underwent enormous changes during the Paleogene as the climate shifted from a greenhouse world towards today’s icehouse, with Paleocene uplift of the Greenland Mantle Plume dramatically changing the paleogeography, oceanography and biotas.
The Early Paleocene (Danian) was characterised by open oceanic conditions and a greenhouse climate that persisted from the Maastrichtian despite the end-Cretaceous bolide impact that devastated much of the planet’s fauna and flora. Marine biotas, including nanoplankton and foraminifera, flourished across the region, with their calcareous remains forming extensive Danian chalk deposits of the Ekofisk Sequence similar to those of the Late Cretaceous (Figure 1).

This benign environment changed dramatically during the Middle and Late Paleocene (Selandian-Thanetian) Maureen and Lista Sequences due to uplift of the Greenland Mantle Plume. Peak uplift and massive extrusion of subaerial and submarine basalts during the Late Thanetian (Forties, Dornoch and Balder Sequences) separated the North Atlantic region into three marine systems.
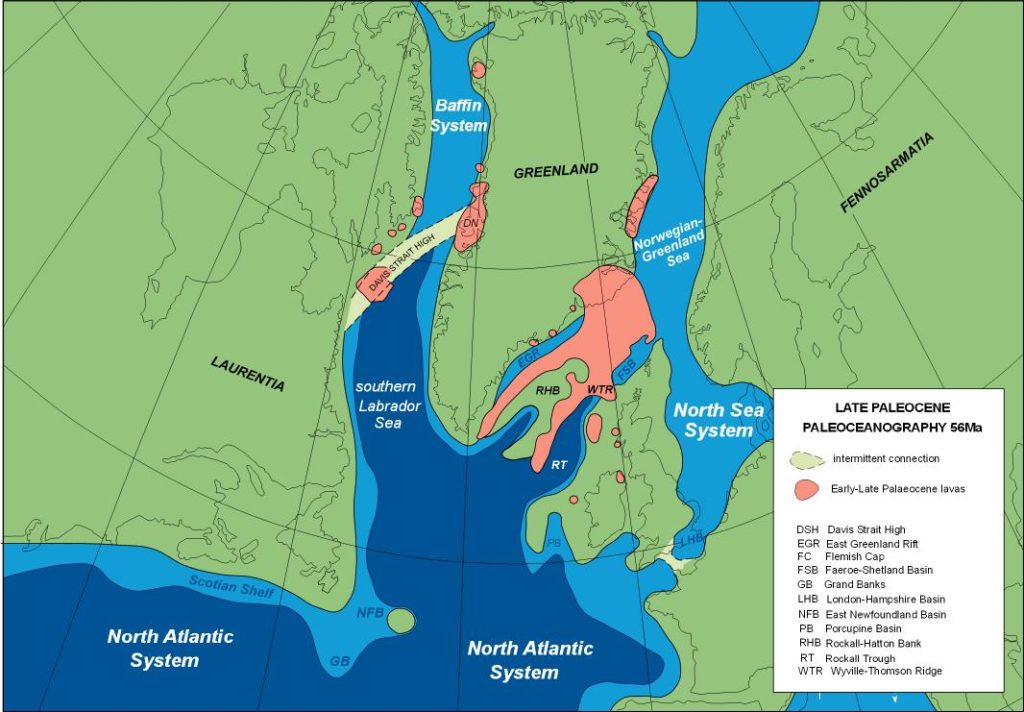
The North Sea System comprised an elongated north-south seaway extending from the London-Hampshire Basin and North Sea northwards to the Norwegian-Greenland Sea (Figure 2). Limited data, primarily from exploration wells indicate that the Baffin System comprised a smaller seaway bounded to the south by the Davis Strait high. In contrast, most of the North Atlantic System, including the Scotian Shelf, Grand Banks, northeast Newfoundland Basin and southern Labrador Sea, had open oceanic conditions with biotas that were more diverse than those of the North Sea and Baffin Systems. The Fareo-Shetland Basin and Rockall Trough formed relatively narrow northeast-southwest seaways of the North Sea and North Atlantic Systems separated by the relatively narrow Wyville-Thomson Ridge, with the Rockall Trough having more diverse assemblages that were similar to those of offshore southeastern Canada.
The oceanographic and tectonic changes had an enormous impact on the sedimentation and biotas of the North Sea and Faroe-Shetland Basin. Calcareous nanoplankton and foraminifera that flourished during the Danian were progressively eliminated as clastic sedimentation increased during the Selandian Maureen Sequence. This altered the water chemistry and microfauna from calcareous-rich assemblages to ones dominated by arenaceous benthic foraminifera and siliceous planktonic radiolaria in the Lista Sequence. Carnivorous dinoflagellates, including Palaeoperidinium, also became more common, supplementing populations that had been dominated by photosynthetic dinoflagellates in the Danian (Figure 3).

As mantle plume uplift reached its peak during deposition of the Late Thanetian and early Ypresian Forties, Dornoch and Balder Sequences, the few remaining benthic foraminiferal species were eliminated from most of the North Sea and Fareo-Shetland Basin due to water stratification and bottom-water anoxia during the North Sea biotic crisis. Marine plankton became increasingly restricted and, by the end of the Thanetian, they were dominated by coscinodiscid diatoms and low-diversity dinoflagellates including carnivorous species of Apectodinium, Cerodinium and Deflandrea that fed on the photosynthetic diatoms. This resulted in plankton blooms that caused local oxygen depletion and dead zones, with the organic remains sinking to the sea floor to be preserved as carbon-rich sapropel and potential source rocks depending on their depth of burial and local geothermal gradient (Figure 3). This is discussed in more detail in next week’s blog.
paleocene – early eocene climate and hyperthermals
The Paleocene and Early Eocene warm greenhouse climate was punctuated by a succession of hyperthermal (HT) events that were triggered by the expulsion of greenhouse gases (GHG) into the atmosphere (Figure 4). These may have been related to northern North Atlantic volcanism, including Selandian and Thanetian volcanics of the east Greenland (Kangerlussuaq) Vandfaldsdalenn and Mikis Formations (e.g., White & Lovell 1997), Thanetian Lower Rockall lavas penetrated by northeast Rockall well 163/25-2 (Bujak, pers. obs.) and the PETM discussed below. Each of the HTs resulted in a temporary influx of warm-water dinoflagellates from the Tethys into the North Atlantic and North Sea.

The earliest HT (WD1) resulted from the Yucatán bolide impact and followed cooling that lasted ‘several decades at most’ when solar radiation was reduced by dust and sulphur aerosols ejected by the impact (Vellekoop et al. 2014).
Increased levels of the greenhouse gas carbon dioxide (CO2) from vaporised carbonate rocks then raised air and sea-surface temperatures for about 100,000 years (Coccioni et al. 2010), enabling the dinoflagellates Manumiella and Trithyrodinium to migrate into higher latitudes (Figure 4). This corresponds to the Dan-C2 global HT listed in Arreguín-Rodríguez et al. (2021).
WD1 was followed by four Middle to Late Paleocene (Selandian-Thanetian) HTs when the warm-water dinoflagellate Apectodinium migrated from the Tethys into mid-latitudes, and the earliest Eocene PETM when its global migration included the Arctic (Figure 4). All of these ‘dinoflagellate migration HTs’ correspond to North Sea / Faroe-Shetland sequence boundaries and four equate with global HTs listed by Arreguín-Rodríguez et al. (2021).
The youngest and most pronounced Apectodinium influx occurred during the Paleocene Eocene Thermal Maximum (PETM) (Bujak & Brinkhuis (1998). The PETM is also called the Early Eocene Thermal Maximum following redefinition of the Paleocene-Eocene Stage boundary from the top to the base of the PETM (see Gradstein et al., 2020). The relevance of this to North Sea and Faroe-Shetland oceanography and petroleum potential is discussed in the second blog in this series.
the eocene: from greenhouse towards icehouse
Greenland Mantle Plume collapse occurred in the earliest Early Eocene (Ypresian), with widespread (London Clay) transgression re-establishing limited Atlantic – North Sea connections that increased during the later Eocene and Oligocene (Figure 5).

The greenhouse world then shifted towards icehouse following the Arctic Azolla Event when large areas of the mainly enclosed Arctic Ocean were repeatedly covered by the freshwater floating fern Azolla that migrated along extensive surface freshwater plumes resulting from high temperatures, rainfall and river discharge (Barke et al. 2011, Brinkhuis et al. 2006, Bujak & Bujak 2021, van Kempen 2013).
The Early Eocene and its greenhouse climate lasted for approximately eight million years, followed by the Arctic Azolla Event (49.3-48.1Ma) which reduced atmospheric CO2 by at least 195 ppm based on calculations of azolla’s Arctic Ocean biomass during the event (Bujak & Bujak 2020, Speelman et al. 2009), similar to values indicated by CO2 proxies (Rae et al. 2021).
The Middle and Late Eocene cooling affected different regions of the North Atlantic due to disconnection of oceanic currents and their heat transfer. The North Sea and Baffin Systems experienced cooler water conditions than the Atlantic System during the Middle and Late Eocene, so that temperature-sensitive dinoflagellate species have different stratigraphic ranges, even in the Rockall and Faroe-Shetland basins that were just a few kilometres apart (Figure 6).

The Terminal Eocene Event resulted from a sudden and dramatic fall in temperature that affected lower latitudes including the Tethys, with the resulting ‘Grande Coupure’ reflecting large-scale extinction and floral and faunal turnover at the beginning of the Oligocene (Van Couvering et al. 1981).
Subsequent oceanic and tectonic processes reduced atmospheric CO2, with brief periods of CO2 increase and warming in the Middle Miocene and Pliocene. Antarctic and Arctic glaciation began approximately 33.7 and 2.6 million years ago (Pagani et al. 2011) leading to today’s bipolar icehouse world. The pre-industrial value of 280 ppm has now increased to 417 ppm, the highest since the Pliocene when there was no Arctic Ocean ice.
Our next blog will discuss the significance of the paleoceanography and climate for energy exploration.
references
Arreguín-Rodríguez GJ et al. 2021. Benthic foraminiferal turnover across the Dan-C2 event in the eastern South Atlantic Ocean (ODP Site 1262). Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 572, ISSN 0031-0182.
Barke J, et al. 2011. Orbitally forced Azolla blooms and Middle Eocene Arctic hydrology: Clues from palynology. Geology. 39(5):427–430. doi:10.1130/g31640.1.
Berggren WA et al. 1995. A revised Cenozoic geochronology and chronostratigraphy. In: Geochronology, Time Scales and Global Stratigraphic Correlations: a Unified Temporal Framework for an Historical Geology. (eds. Berggren, W.A., Kent, D.V., Aubrey, M.-P. & Hardenbol, J.). Society of Economic Palaeontologists and Mineralogists Special Publication, 54, pp. 129-212.
Brinkhuis H, et al. 2006. Episodic fresh surface waters in the Eocene Arctic Ocean. Nature. 441(7093):606–609. doi:10.1038/nature04692.
Bujak JP 2023. Palaeocene-Eocene hydrocarbon potential of the Faroe-Shetland Basin based on high-resolution stratigraphy, oceanography and climate. Non-exclusive report.
Bujak JP & Bujak AA. 2021. The Azolla Story. Published by the Azolla Foundation, available as an e-book and print version: https://theazollastory.com/
Bujak, JP & Mudge DC 1994. A high-resolution North Sea Palaeocene – Eocene dinocyst zonation. Journal of the Geological Society of London, vol. 151: 449-462.
Coccioni R et al. 2010. The Dan-C2 hyperthermal event at Gubbio (Italy): Global implications, environmental effects, and cause(s). Earth and Planetary Science Letters, vol. 297(1-2), pp. 298-305.
Gradstein FM et al. 2020. Geologic time scale 2020, 2nd Edition, 1 March 2021. Amsterdam: Elsevier.
Kempen M van. 2013. Azolla on Top of the World: an Ecophysiological Study of Floating Fairy Moss and Its Potential Role in Ecosystem Services Related to Climate Change. Darwin Centre for Biogeosciences, Radbiud University Nijmegen.
Meinshausen M et al. 2019. The SSP greenhouse gas concentrations and their extensions to 2500. Geosci. Model. Dev. Discuss: 1–77.
Mudge DC & Bujak JP. 1996. An integrated stratigraphy for the Paleocene and Eocene of the North Sea. In: Correlation of the Early Paleogene in Northwest Europe (Eds Knox et al.). Geological Society Special Publication No. 101(1): 91-113
Pagani M et al. 2011. The Role of Carbon Dioxide During the Onset of Antarctic Glaciation. Science, vol. 334, issue 6060: 1261-1264.
Pearson PN & Palmer MR. 2000. Atmospheric carbon dioxide concentrations over the past 60 million years. Nature. 406(6797): 695–699.
Rae JWB et al. 2001. Atmospheric CO2 over the Past 66 million Years from marine archives. Annual Review of Earth and Planetary Sciences, vol. 49: 599–631.
Van Couvering JA et al. 1981. The Terminal Eocene Event and the Polish connection. Palaeogeography, Palaeoclimatology, Palaeoecology, vol 36(3–4): 321-362.
Vellekoop J et al. 2014. Earth, Atmospheric and Planetary Sciences, vol. 111(21): 7537-7541 https://doi.org/10.1073/pnas.1319253111 Rapid short-term cooling following the Chicxulub impact at the Cretaceous–Paleogene boundary.
White N & Lovell B. 1997. Measuring the pulse of a plume with the sedimentary record. Nature, vol. 387: 888-891.
Zachos JC et al. 2003. A transient Rise in Tropical Sea Surface Temperature During the Paleocene-Eocene Thermal Maximum. Science Reports, vol. 302: 1551-1554.
Zachos JC et al. 2005. Rapid Acidification of the Ocean During the Paleocene-Eocene Thermal Maximum. Science, vol. 308(5728): 1611-1615.



{kind=link}